Автор: В. В. Гриднева
ДИНАМИКА ЭКОСИСТЕМ И ИХ КОМПОНЕНТОВ
УДК 630*182.21/574.42 /598.2
Ивановский государственный университет
Россия, 153002, г. Иваново, просп. Ленина, д. 136. E-mail: gridnevavv@mail.ru
Смена бореальных сообществ вторичными мелколиственными лесами в результате переэксплуатации в условиях существующего подхода к ведению лесного хозяйства характеризует большинство подтаежных лесных орнитоценозов Восточного Верхневолжья. Оценка уже имеющихся и возможных последствий этих смен имеет как прикладное так и теоретическое значение.
На основе картографических площадочных учётов птиц в модельных сукцессионных рядах прослежена динамика сообществ. Модельные ряды охватывают все применяемые способы рубок в разных для типах лесов региона. Проведен факторный анализ плотности гнездования, показателей сходства населения птиц на модельных площадках и структуры фитоценоза обследованных местообитаний.
В статье представлены топические предпочтения отдельных видов птиц, использующих сукцессионные биотопы в эксплуатируемых лесах, ключевые характеристики трансформированных рубками местообитаний, определяющие видовой состав, плотность населения и структуру орнитоценоза. Приведена схема ординации сообществ птиц в эксплуатируемых лесах Восточного Верхневолжья методом многомерного неметрического шкалирования. Прослежена направленность сукцессионных антроподинамических изменений сообществ лесных птиц в сторону бореальных сообществ, при почти полной невозможности достижения сообществом климаксных стадий в условиях современных норм ведения лесного хозяйства.
По результатам исследования возможна разработка рекомендаций к уходу за лесами на конкретных территориях региона с учётом гнездования обычных и редких видов птиц. Ключевые слова: рубки леса, сукцессия, динамика населения птиц, Восточное Верхневолжье. DOI: 10.24411/2542-2006-2018-10016
В настоящее время в Восточном Верхневолжье ведется комплексная работа по изучению динамики авифауны в ходе демутационных посттехногенных сукцессий различных экосистем (Мельников, 2010), и данные об антроподинамических восстановительных сукцессиях лесной авифауны после рубок значительно дополняют проведенные ранее исследования.
Лесами занята почти половина территории Восточного Вехневолжья, и эксплуатация леса — наиболее распространённый вид техногенной трансформации, а сукцессии, происходящие на вырубках и гарях, остаются самой масштабной формой современной антропогенной динамики естественных сообществ как в изучаемом регионе, так и в целом по нашей стране. Регион исследования находится на южной границе бореальной зоны, и из-за расположения в самом центре Русской равнины все экосистемы на протяжении длительного времени подвержены значительной антропогенной трансформации. Смена бореальных сообществ вторичными мелколиственными лесами в результате переэксплуатации в условиях существующего подхода к ведению лесного хозяйства характерна для данных территорий (Ярошенко, 1999). Изучение сукцессионных смен на техногенно трансформированных территориях, переведенных в лесопокрытую площадь и, согласно современным нормам ведения лесного хозяйства, развивающихся с постоянным вмешательством, даст возможность оценить, насколько катастрофичны последствия смены породного состава лесов для современной авифауны.
Основной целью данной работы является прогноз тенденций динамики населения птиц в ходе сукцессионных изменений, вызванных лесоэксплуатацией и уходом за лесами в Восточном Верхневолжье. Для достижения цели были поставлены следующие задачи.
1. Подбор биотопов в разных типах эксплуатируемых лесов региона, измененных характерными способами рубки и последующими сукцессионными процессами и балльная оценка основных структурных характеристик этих биотопов.
2. Определение видового состава и показателей структуры населения птиц в подобранных местообитаниях.
3. Выявление ключевых топических факторов, определяющих структуру авифауны на лесных территориях, измененных рубками.
4. Определение трендов динамики орнитоценозов Восточного Верхневолжья в ходе сукцессионных изменений, опосредованных лесохозяйственной деятельностью.
Материалы и методы
Исследуемые территории. Восточное Верхневолжье — регион в центре нечерноземной зоны Русской равнины в бассейне реки Волги в пределах Горьковского водохранилища и среднего течения реки Клязьмы. Он сильно мозаичен и имеет сложную структуру растительности. Лесами занято около 40% территории. С северо-запада на юго-восток проходит граница двух лесных подзон — южной тайги и смешанных лесов. Основная работа велась на 4 стационарах (рис. 1), расположенных в юго-западной, центральной и северной частях региона. Фаунистический состав населения птиц измененных лесоэксплуатацией территорий уточнялся в ходе обследования дополнительных лесных массивов на неохваченных стационарными исследованиями территориях.
Лесопользование, в том числе и лесоэксплуатация, в пределах изучаемых стационаров на протяжении нескольких десятилетий постоянно поддерживаются на среднем уровне интенсивности. Эксплуатируемые леса на территории стационаров являются наиболее типичными для Восточного Верхневолжья — это ельники, смешанные хвойно-мелколиственные леса, а также сосняки-беломошники и зеленомошники. В них применяются как сплошнолесосечные рубки, так и выборочные. Среди рубок ухода наиболее распространены проходные рубки, в сосняках применяются прореживания, в смешанных лесах — рубки переформирования. В связи с вспышками численности стволовых насекомых и пожарами довольно большие площади занимают санитарные рубки. Наиболее удаленный от густонаселенных районов стационар Красногорский характеризуется отсутствием рубок ухода и санитарных рубок.
Исследования проводились в сообществах, сформированных в ходе сукцессий после рубок разных целей и технологий, которые характерны как для территории стационаров, так и в целом для региона. Для каждого типа эксплуатируемых лесов подбирался модельный лесной массив, где закладывался ряд площадок на различной степени сукцессионного зарастания, которые условно объединялись в серию — это открытые вырубки, закустаренные вырубки и жердняки. Дальнейший ход сукцессионных изменений фауны и населения птиц лесных территорий после смыкания древостоев на вырубках и гарях, связан с изменениями, опосредованными уходом за лесными культурами, а также с эксплуатацией лесов выборочными рубками. Эти процессы (воздействие частичных рубок на вторичные леса) мы отслеживали, закладывая площадки как в последующий рубке гнездовой сезон, так и по прошествии сообществом заметных для населения птиц скуцессионных изменений -восстановления лесной подстилки и нижних ярусов.
Рис. 1. Расположение стационаров на территории Восточного Верхневолжья. Fig. 1. Locations of stations on the territory of the Eastern Upper Volga Region.
ЭКОСИСТЕМЫ: ЭКОЛОГИЯ И ДИНАМИКА, 2018, том 2, № 3
Методы исследования. Основным методом являлся абсолютный учёт птиц с картированием гнездовых территорий на пробных площадках (Гудина, 1999), за пробную площадку принималась вся территория, подвергшаяся рубке, или (реже) ее часть в случае больших площадей у пройденных санитарными рубками гарей и поврежденных стволовыми вредителями лесов. Площадки, относящиеся в исследовании к одной сукцессионной серии, обследовались в один год, чтоб нивелировать влияние сезонных флуктуаций численности и погодных условий. Картирование гнездовых территорий сопровождалось бальной оценкой основных структурных топических характеристик сообщества и геоботаническим описанием, на основе которых затем проводился корреляционный анализ (коэффициент ранговой корреляции Спирмена), помогавший при интерпретации результатов статистических расчетов.
Плотность населения (Ni) гнездящихся пар рассчитывалась на 1 га. Исходя из полученных значений рассчитывалась доля участия вида в населении (Рi) в % и долях от единицы для расчета индекса сходства.
Для сравнения населения птиц обследованных местообитаний использован коэффициент сходства Жаккара, модифицированный Р.Л. Наумовым (1964), расширенный по доминированию (Ii(Рi)):
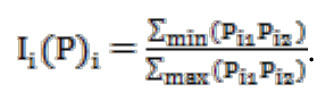
Индекс определяли как отношение суммы минимальных значений индекса доминирования каждого вида из отмеченных в паре сравниваемых сообществ к сумме максимальных значений.
На основе рассчитанных индексов была построена матрица сходств между биотопами и проведено непараметрическое многомерное шкалирование.
Ключевые факторы, определяющие распространение видов в изучаемых биотопах, определяли методом анализа главных компонент без вращения факторных нагрузок.
Статистическая обработка данных проводилась при помощи программного пакета StatSoft STATISTICA 6.1.478 Russian 2007.
Русские и латинские названия птиц и их систематика в работе даны по сводке Е.А. Коблика с соавторами (2006). Достоверность гнездования вида в обследованных биотопах при отсутствии возможности обнаружения гнезда определялась по критериям, рекомендованным изданием «The EBCC Atlas of European Breeding Birds» (1997).
Материалы исследования. Основная часть работы основана на данных учётов птиц, проведенных в весеннее-летние сезоны с 2006 по 2010 год (табл. 1). Общая площадь обследованных площадочным методом биотопов составила 324 га, которые пройдены многолетними и многократными картографическими учётами. Представленный ниже анализ основывается на выявленных в ходе картографирования 461 гнездовых участков 46 видов птиц. Стоит пояснить, что выявленная орнитофауна трансформированных рубками территорий, не охваченных площадочным учётом, значительно обширней, сукцессионные биотопы на месте рубок в репродуктивный период используют 67 гнездящихся видов, 5 -возможно, гнездится, 12 используют восстанавливающиеся после рубок лесные территории как кормовые стации.
Результаты исследования и их обсуждение
В ходе исследования выявлено 86 видов птиц, использующих измененные рубками лесные территории Восточного Верхневолжья и относящихся к 12 отрядам, что составляет 65% от гнездящихся в регионе лесных видов (Мельников и др., 2005). Гнездовая авифауна насчитывает 73 вида, большинство из которых — обычные лесоопушечные и толерантные лесные виды. Спектр доминантов довольно широк, — насчитывается 7 фоновых видов преобразованных рубками территорий, каждый из которых доминирует в группе биотопов определенного этапа сукцессии. Видовой состав птиц, использующих лесные территории, подвергавшиеся рубкам, имеет отличия в лесах разных типов. На него влияют преобразованных рубками лесных территорий как зонально-подзональные отличия авифауны, так и характер ведения заготовительных и лесовосстановительных работ, большую роль играет пирогенный фактор.
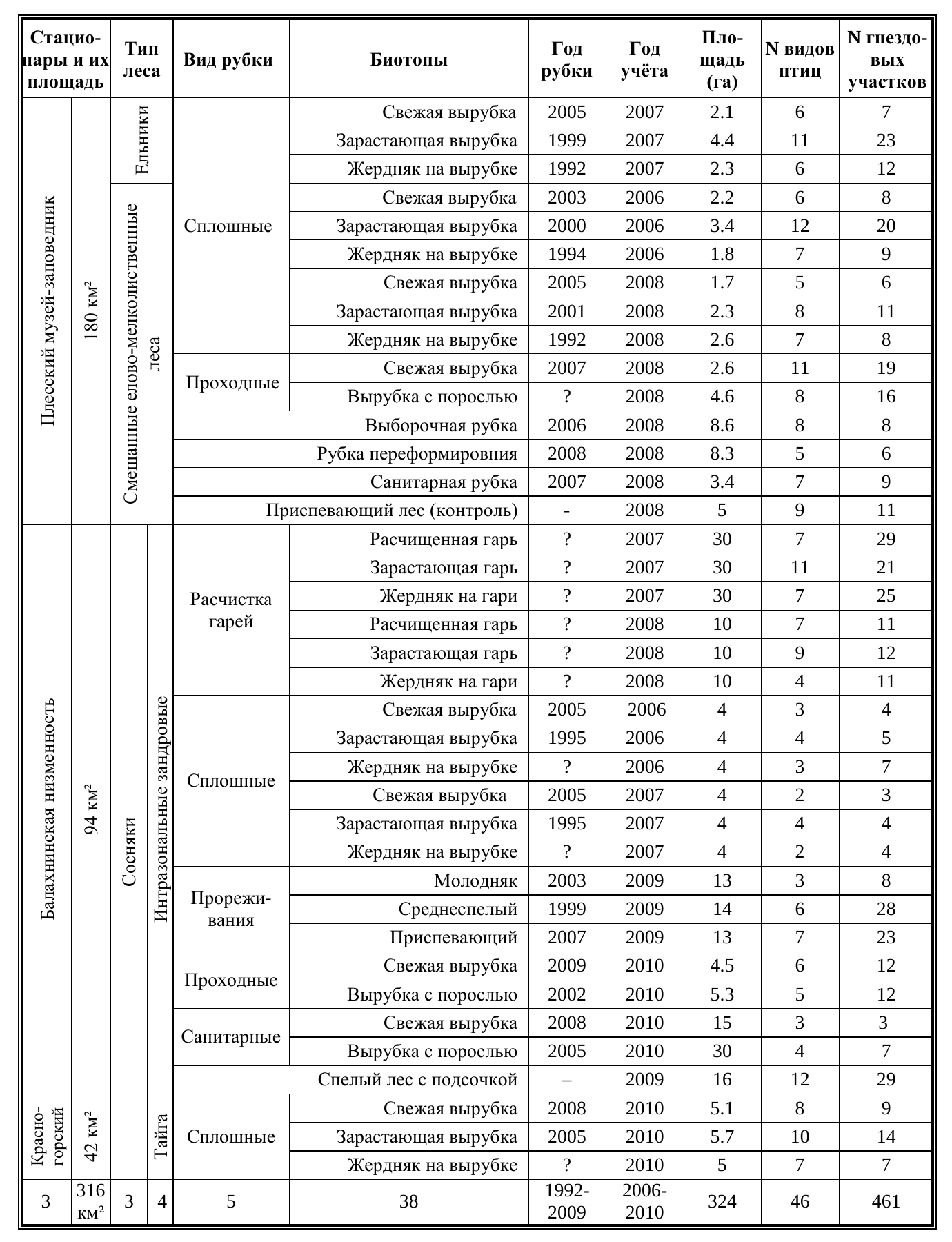
Таблица 1. Материалы площадочных учётов, использованные в работе. Table 1. Data of site census, used in our work.
При анализе зависимостей показателей численности отдельных видов птиц с бальным выражением структурных характеристик биотопов было выявлено несколько значимых на среднем уровне корреляций (табл. 2) с такими топическими характеристиками как площадь вырубленного пространства, площадь открытого пространства, протяженность экотонных территорий, мозаичность, сомкнутость крон, ярусность, заболоченность и наличие выгоревших участков. Также выявлены корреляции плотности населения для ряда видов с распространением определенных растительных формаций. Так, например, наличие мелколиственного подроста привлекает на гнездование козодоя и весничку. Старовозрастные деревья — важный элемент гнездового биотопа пеночки-теньковки. Следует особо отметить, что большинство указанных видов демонстрируют слабый или средний уровень коррелляции.
Анализируя плотность населения видов в рассматриваемых биотопах методом главных компонент (факторный анализ), удалось выявить ключевые факторы, влияющие на распространение и численность большинства видов на пройденных рубками территориях (рис. 2). Это, во-первых, степень трансформации биотопа в ходе рубки (нарушение древесного полога и лесной подстилки), причем полная нарушенность в ходе сплошных рубок противопоставляется частичной, являющейся результатом рубок ухода. Во-вторых, это мозаичность биотопа, в-третьих — его открытость (параметр, обратный сомкнутости крон).
В пространстве ключевых биотопических факторов выделяется несколько группировок видов со сходными требованиями, которые подтверждают результаты, полученные при анализе корреляций численности видов с выделенными в ходе обследования биотопов структурными характеристиками. Это группа видов открытых пространств, группа опушечных видов, группа видов, предпочитающих разреженные леса и молодняки, и группа толерантных к рубке лесных видов.
Так, перепел (Coturnix coturnix) предпочитает низкомозаичные открытые биотопы, восстанавливающиеся после сплошных рубок, крапивник (Troglodytes troglodytes) — сходные, но менее открытые. Белая трясогузка (Motacilla alba) и чечевица (Carpodacus erithritnus) способны гнездиться при сильном нарушении древостоя и целостности лесной подстилки в ходе сплошных рубок, также предпочитая маломозаичные биотопы. Бекас (Gallinago gallinago), жёлтая трясогузка (Motacilla flava), обыкновенная овсянка (Emberiza citrinella) и луговой чекан (Saxicola rubetra) занимают максимально открытые маломозаичные сплошные вырубки, на которых только начинается восстановление почвенного покрова и древостоя. Большой улит (Tringa nebularia), полевой жаворонок (Alauda arvensis), обыкновенная каменка (Oenanthe oenanthe) и юла (Lullula arborea) населяют менее открытые и нарушенные биотопы, соответствующие вейниковым пустошам на гарях. Перечисленные виды относятся к группе видов открытых пространств.
Садовая славка (Sylvia borin), обыкновенный соловей (Luscinia luscinia), садовая камышевка (Acrocephalus dumetorum) и дрозд белобровик (Turdus iliacus) населяют полуоткрытые высокомозаичные биотопы, частично восстановившие нарушение лесной подстилки после сплошных рубок и имеющие древесный подрост.
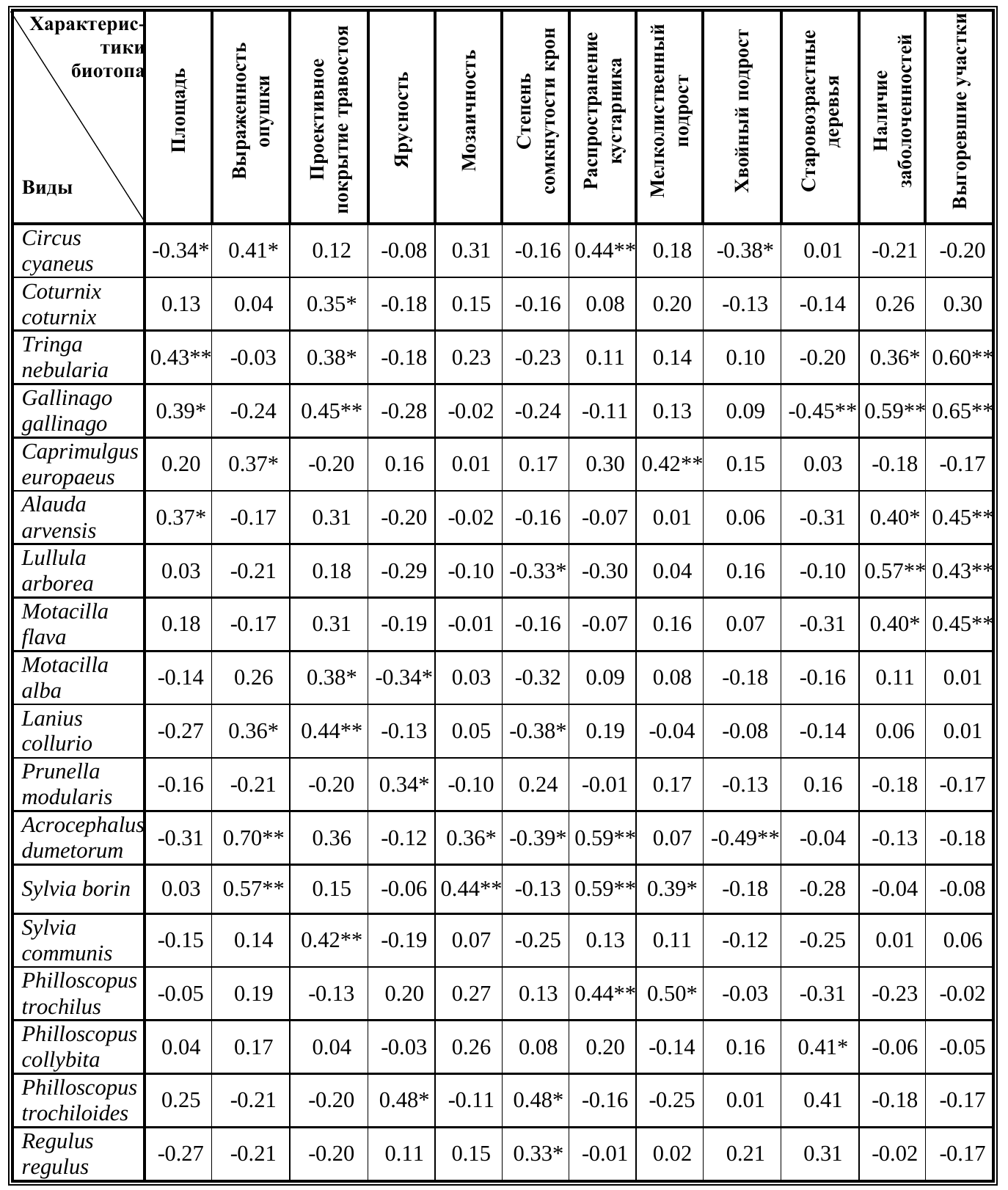
Таблица 2. Значения коэффициентов корреляции Спирмена между плотностью гнездования и структурными характеристиками биотопа. Table 2. Values Spearman&s rank correlation coefficient between nesting density and structural features of biotope.
Примечания к таблице 2: * — значимы на уровне P≤0.05, ** — значимы на уровне P<0.01. Notes to table 2: * — values are significant at P≤0.05, ** — values are significant at P<0.01.
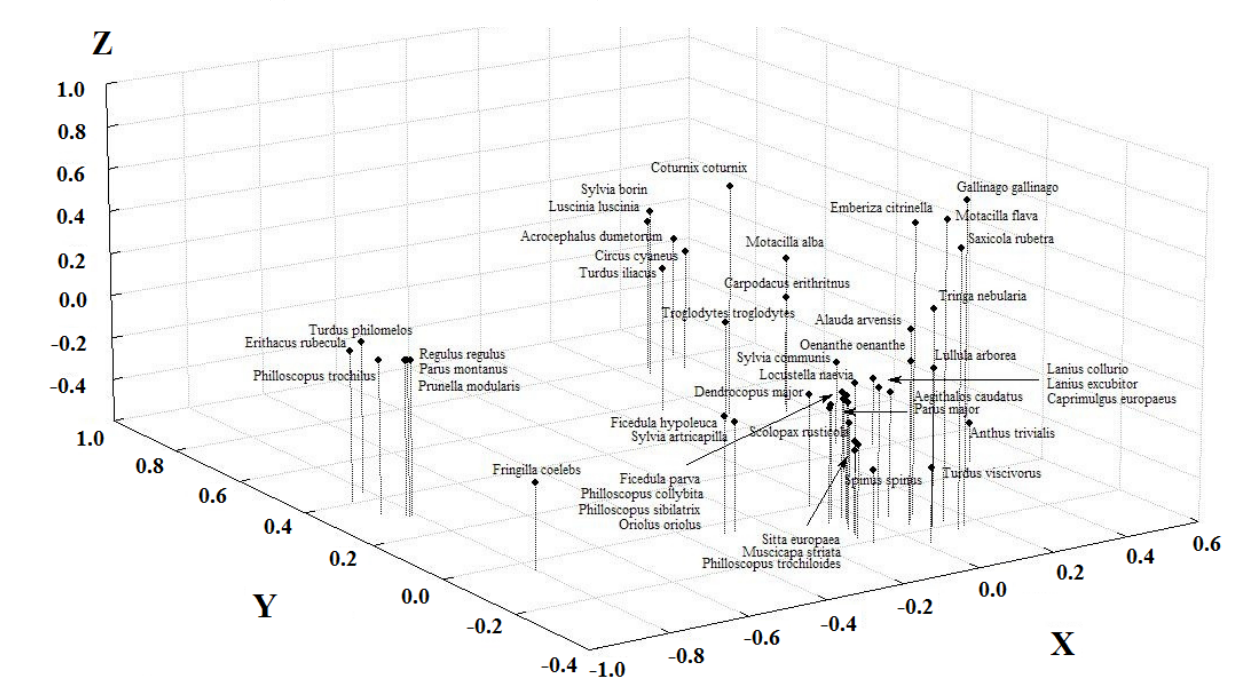
Рис. 2. Распределение гнездящихся в эксплуатируемых лесах видов птиц в пространстве ключевых биотопических факторов. Условные обозначения: ось Х — деструкция полога леса и лесной подстилки (последствия частичных рубок откладываются на оси в отрицательную сторону, противопоставляясь сплошнолесосечным); ось Y — мозаичность; ось Z — открытость (параметр, обратный сомкнутости крон в лесу). Fig. 2. Distribution of birds’ species, nesting in the exploited forests in the area of key biotopical factors. Legend: Х — destruction of forest canopy and cover (aftermath of partial felling are plotted on the negative direction, opposed to clear-cutting); Y — patchiness; Z — openness (which is opposed to canopy density).

Серый сорокопут (Lanius excubitor) и обыкновенный жулан (Lanius collurio) занимают нишу маломозаичных восстанавливающихся после сплошных рубок полуоткрытых лесных территорий на гарях и вырубках. Серая славка (Sylvia communis) и обыкновенный сверчок (Locustella naevia) занимают сходные биотопы с большей мозаичностью и относятся к опушечным видам.
Певчий дрозд (Turdus philomelos), зарянка (Erithacus rubecula) и пеночка-весничка (Philloscopus trochilus) предпочитают маломозаичные биотопы с отсутствием сомкнутости крон и минимумом открытых пространств при частичном нарушении лесной подстилки вследствие рубок ухода. Жёлтоголовый королек (Regulus regulus), буроголовая гаичка (Parus montanus) и лесная завирушка (Prunella modularis) предпочитают сходные биотопы с еще меньшей мозаичностью.
Мухоловка-пеструшка (Ficedula hypoleuca) и славка-черноголовка (Sylvia artricapilla) способны гнездится в малонарушенных после частичных рубок лесах с низкой сомкнутостью крон и отсутствием мозаичности. Зяблик (Fringilla coelebs) может населять подобные биотопы и при сильном нарушении в ходе частичных рубок. Монотонные лесные биотопы, лишенные мозаичной структуры используются для гнездования лесным коньком (Anthus trivialis) и дерябой (Turdus viscivorus). Эти виды относятся к группировке птиц молодняков и разреженных лесов.
Остальные виды населяют леса поздних этапов восстановления после сплошных или частичных рубок с формирующейся сомкнутостью крон и низкой мозаичностью. Эти виды характерны и для ненарушенных рубками старовозрастных, и для вторичных спелых лесов.
Хотя перестройки авифауны в ходе рубок и последующей сукцессии лесов являются исторически сложившимися и чаще не катастрофичны, большинство видов птиц, использующих измененные рубками лесные территории, — это обычные лесоопушечные и толерантные к антропогенной трансформации местообитаний лесные виды. Большинство гнездящихся редких видов (в частности, занесенных в региональные Красные Книги), заселяя вырубленные территории, требуют наличия специфических условий, таких как переувлажненные местообитания; характерные для вырубок микроместообитания в завалах порубочных остатков, фаутные 1 и семенные деревья. Редкими видами птиц, способными заселять непосредственно вырубки, являются лесной жаворонок (Lullula arborea) и ястребиная славка (Sylvia nisoria). Змееяд (Circaetus gallicus), большой подорлик (Aqila clanga), беркут (Aqila chrysaetos) и филин (Bubo bubo) используют вырубки и гари как охотничьи территории. Полевой лунь при выборе гнездовых участков демонстрирует предпочтение вырубленных лесных территорий, при этом наблюдается его переход с внешних опушек леса на внутренние. Смену стереотипа гнездования в измененных рубками биотопах демонстрируют и другие виды (обыкновенный жулан, крапивник, обыкновенная каменка).
1 Фаутные деревья (англ. faut — дефект) — деревья с наличием пороков ствола (фаутов): дупел, ошмыгов, сухобокости, трещин, отлубов, наплывов … (прим. ред.).
Количество гнездящихся видов, для которых оптимальны сложившиеся после рубки или в ходе сукцессии условия, редко выходит за пределы в 5-S (табл. 1). Следует отметить, что на начальных стадиях постпирогенной сукцессии гнездится на 1 -2 вида больше, чем в биотопах, сформированных рубками в незатронутых пожарами лесах. Причем, видов, использующих для гнездования исключительно пирогенные биотопы, не выявлено, — все гнездящиеся на расчищенных гарях виды делают это и в биотопах, сформированных рубками в отсутствие пирогенного воздействия. Присутствие сравнительно большего спектра видов объясняется возможностью сосуществования видов с разными требованиями к размерам индивидуальных территорий в пределах открытых биотопов большой площади, лишенных остатков предыдущего сообщества полностью. Максимальный показатель видового богатства характерен для мозаичных биотопов, сформированных на месте сплошных вырубок и гарей после разрастания кустарников и древесного подроста — 11-12 видов, а также старовозрастных вторичных лесов с выраженной ярусностью.
Низкие показатели количества гнездящихся видов были отмечены на свежих вырубках сосновых лесов Балахнинской низины и в молодых хвойных посадках (2-4 вида). Малое количество видов, гнездящихся на открытых и зарастающих вырубках соснового леса, по-видимому, обусловлено ограниченной площадью биотопа и его монотонностью. При этом вырубки сосняков на территории Красногорского стационара, не существенно превышающие по площади вышеописанные, характеризуются большим количеством видов, близким к таковому на вырубках смешанных лесов и ельников. Эти различия объясняются искусственным лесовозобновлением после рубок, отсутствие которого для любого типа лесов региона чревато перенаправлением сукцессионного процесса в сторону вторичных хвойно-мелколиственных лесов.
На основе матрицы сходства обследованных биотопов было проведено многомерное шкалирование (рис. 3), по результатам которого в двумерном пространстве определены дистанции между показателями сходства всех обследованных биотопов по обилию гнездящихся видов птиц (индекс сходства Жаккара, расширенный по доминированию). При интерпретации полученных осей выявили, что близость населения птиц сформированных различными типами рубок и последующей сукцессией биотопов обусловлена обилием массовых видов птиц, приуроченных преимущественно к начальным этапам сукцессии лесов после различных способов рубки, а также сходными показателями обилия видов, характерных для финальных стадий сукцессионного восстановления (отдельно для лиственных и хвойных лесов).
Результаты ординации орнитокомплексов позволяют выделить в качестве исходных точек начала антроподинамических сукцессий группы орнитокомплексов недавно пройденных сплошной рубкой лесов различных типов (смешанные леса и ельники, сосняки различного происхождения и гари соснового леса). На имеющейся выборке орнитокомплексов прослеживается общая направленность сукцессионных смен от далеко отстоящих друг от друга сообществ открытых вырубок в разных типах лесов к большой группе сходных сообществ птиц молодых вторичных лесов на разной степени спелости с незначительными отличиями, привносимыми рубками ухода.
Следует отметить, что вся довольно разнообразная по технологии рубок и исходному типу леса выборка трансформированных территорий характеризуется сравнительно высокими показателями сходства населения птиц на промежуточных этапах сукцессии.
Полученное распределение позволяет говорить о возможной направленности антроподинамических сукцессионных изменений в населении птиц лесных экосистем Восточного Верхневолжья в сторону сообществ разновозрастных хвойных лесов близких к естественным бореальным. Такой исход сукцессионных смен возможен при неукоснительном соблюдении норм ведения лесного хозяйства (утопичном в современных условиях для данной территории). Наиболее близки по фауне и населению к южнотаежным сообщества птиц спелых многократно прореженных сосняков-беломошников и пройденных рубками переформирования (удалением лиственных пород) смешанных лесов. Применение рубок ухода, таких как постепенные прореживания, проходные рубки и рубки переформирования, формирует в сложной мозаике современных разновозрастных вторичных лесов небольшие выделы многоярусных хвойных насаждений, которые способны поддерживать сообщества с типично-таежными видами птиц, редкими для остальных участков эксплуатируемых лесных массивов. Развивающиеся естественным путем, в отсутствии искусственного лесовозобновления и мероприятий по уходу за лесонасаждениями территории, являясь малопривлекательными для эксплуатации, по достижении сообществом зрелости поддерживают сообщества птиц хвойно-мелколиственных, а в отсутствии катастрофических нарушений длительное время (в труднодоступных для транспорта районах) и хвойно-широколиственных старовозрастных лесов.
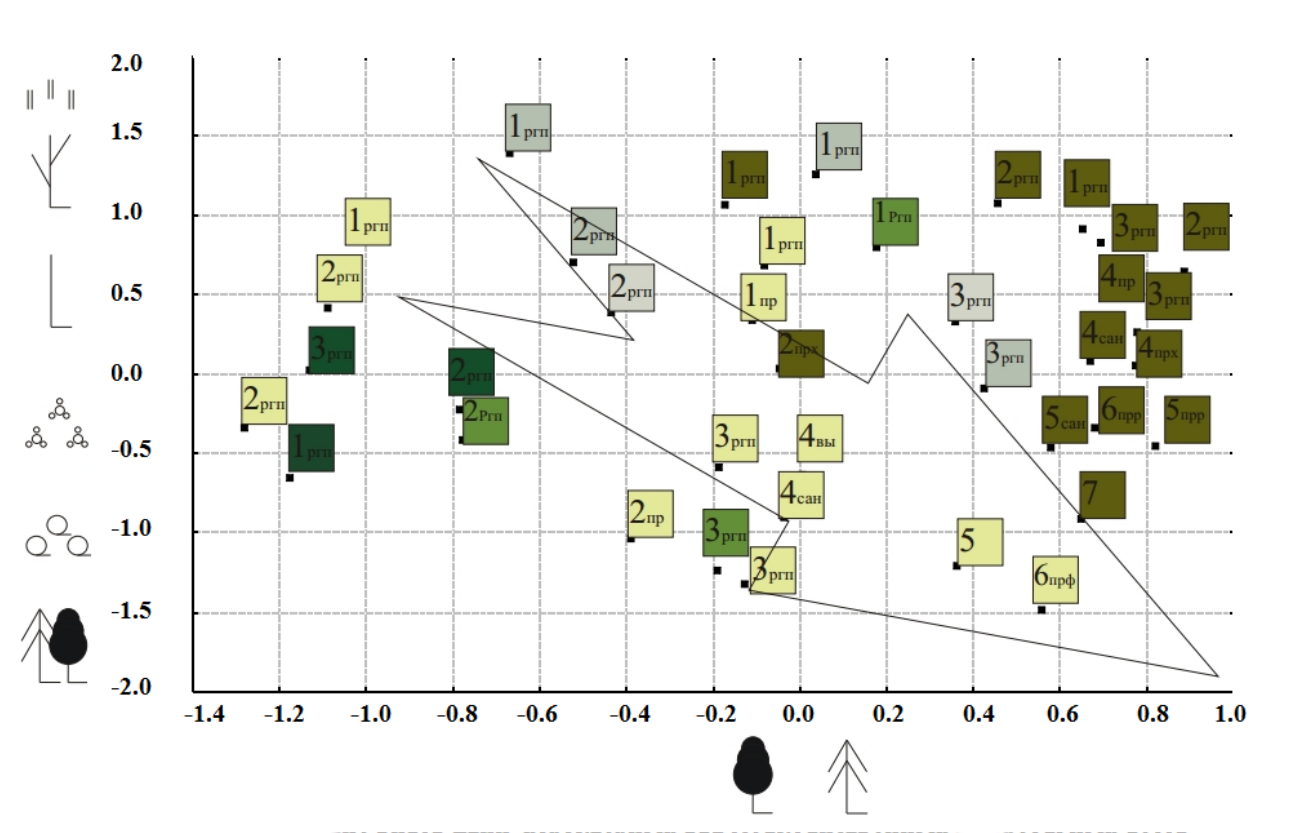
Рис. 3. Экологическая ординация орнитокомплексов сформированных сукцессией после рубок. Условные обозначения. По вертикальной оси: обилие видов птиц, характерных для

лугов и пустошеи,
— пирогенных стации,

— недавних лесозаготовок,
— опушек и пойм,
— разреженных лесов и буреломов,

— сомкнутых древостоев со старовозрастными деревьями.
По горизонтальной оси:

— обилие видов птиц, характерных для мелколиственных лесов,
— обилие бореальных видов птиц.
Цветом квадратов обозначено предшествующее рубке сообщество:

— гари сосняков-беломошников,


-сосняки-беломошники,

— подтаежные сосняки,

— смешанные хвойно-мелколиственные леса,
-ельники.
Цифрами в квадратах (1, 2, 3 …) обозначены условные этапы сукцессии после рубки: спл. — сплошнолесосечные рубки, сан. — санитарные рубки поврежденных пожарами или стволовыми вредителями лесов, прх. — проходные рубки, прр. — прореживания в хвойных посадках разного возраста, прф. — рубки переформирования изъятием лиственных пород, выб. — выборочные рубки привлекательных стволов. Стрелка показывает тренд антроподинамической сукцессии.
Fig. 3. Ecological ordination of ornithocomplexes, formed by succession after forest felling. Legend. Along the vertical axis: birds& species richness, specific for
— meadows and wastelands,
— pyrogenic stations,
— recent forest harvesting,
— forest edges and floodplains,
— broken forests and windfalls,
— closed forests with old trees.
Along the horizontal axis:
— birds’ species richness, specific for small-leaved forests,
— boreal birds’ species richness.
The squares stand for communities, preceding the felling:
— burned areas of lichen pine forests,
— lichen pine forests,
— subtaiga pine forests,
— mixed coniferous-small-leaved forests,
— spruce forests.
The numbers in the squares (1, 2, 3 etc.) stand for provisional stages of succession after felling: спл. — clear-cutting felling, сан. — sanitary felling of the forests, damaged with fire or vermin insects, прх. — severance felling, прр. — cutting-back of spruce forests of different ages, прф. — felling of reformation by the removal of deciduous species, выб. — random felling of fine trunks. The arrow shows the trend of anthropodynamical succession. транспорта районах) и хвойно-широколиственных старовозрастных лесов.
Современная динамика населения птиц в ходе сукцессионных изменений эксплуатируемых лесов Восточного Верхневолжья с учётом лесохозяйственных мероприятий сводится к 2 типам — это динамика, приводящая к сообществам смешанных, хвойно-мелколиственных лесов, и динамика по типу пирогенных зандровых сосняков. Оба эти типа динамики поддерживаются нормами ведения лесного хозяйства на протяжении длительного времени (более полутора столетий) при стремящейся к нулю вероятности перехода из одного в другой и полной невозможности достижения климакса (сообщества птиц разновозрастного ельника или елово-широколиственного леса).
Выводы
1. Авифауна измененных лесоэксплуатацией территорий насчитывает 86 видов, среди которых немногочисленные регионально редкие приурочены к специфическим микроместообитаниям, формируемым на вырубленных территориях другими факторами.
2. Гнездование ряда видов и их плотность населения зависит от таких специфических характеристик биотопа, измененного рубкой, как площадь, выраженность опушки, а также наличия порубочных остатков, выгоревших участков и распространения определенных растительных формаций.
3. Ключевыми факторами, определяющими распространение птиц на пройденных рубками территориях являются: степень нарушения, мозаичность, открытость биотопа (показатель, обратный сомкнутости крон).
4. Антроподинамические сукцессии орнитокомплексов в лесах Восточного Верхневолжья можно разделить на 2 основных типа — смены населения по типу смешанных лесов (с отсутствием систематического ухода), характеризующиеся высоким разнообразием на начальных этапах, и смены по типу пирогенных сосняков (с систематическим уходом и частой угрозой выгорания, приводящим к распространению одновозрастных моновидовых сосновых культур) со специфической упрощенной структурой населения птиц.
5. Общим трендом сукцессионной динамики орнитоценозов в эксплуатируемых лесах Восточного Верхневолжья с учётом современных лесохозяйственных мероприятий является постепенное снижение доли видов открытых пространств вплоть до полного исчезновения при постепенном вытеснении видов характерных для экотонов и мелколиственных лесов бореальными видами.
СПИСОК ЛИТЕРАТУРЫ
Гудина А.Н. 1999. Методы учёта гнездящихся птиц. Картирование территорий. Запорожье: Дикое поле. 241 с.
Коблик Е.А., Редькин Я.А., Архипов В.Ю. 2006. Список птиц Российской Федерации.
М.: Товарищество научных изданий КМК. 256 с. Мельников В.Н. 2010. Динамика орнитокомплексов в ходе зарастания территорий,
выведенных из хозяйственного использования в условиях Восточного Верхневолжья // Орнитология в Северной Евразии: Материалы XIII Международной орнитологической конференции Северной Евразии / Ред. В.Н. Мельников Оренбург: ОГПУ. С. 217.
Мельников В.Н., Баринов С.Н., Киселев Р.Ю., Чудненко Д.Е. 2005. Современное состояние изученности авифауны Ивановской области // Природа и человек. Материалы IV научно-практической конференции «Природа и человек. Антропогенное воздействие на окружающую среду». Иваново. С. 127-129.
Наумов Р.Л. 1964. Опыт абсолютного учёта лесных певчих птиц в гнездовой период // Организация и методы учёта птиц и вредных грызунов. М.: АН СССР. С. 137-147.
Ярошенко А.Ю. 1999. Европейская тайга на грани тысячелетий. М.: Гринпис России. 66 с.
The EBCC Atlas of European Breeding Birds. 1997. Their Distribution and Abundance. London. 903 р.
DYNAMICS OF BIRDS& POPULATION OF THE EASTERN UPPER VOLGA REGION, MEDIATED BY THE MODERN FOREST EXPLOITATION
© 2018. V.V. Gridneva
Ivanovo State University Russia, 153002, Ivanovo, Leninsky Avenue, 136. E-mail: gridnevavv@mail.ru
Replacement of the boreal communities with secondary small-leaved forests as a result of overexploitation in the conditions of the existing method of forestry management is common for the most subtaiga forest ornithocenosis of the Eastern Upper Volga Region. Evaluation of every existing and future aftermath of this replacement has an applied and theoretical meaning.
The dynamics of the communities are traced on the basis of cartographic sited census in the modelling succession ranges. Model ranges cover all methods of felling in different forests of the region. We made a factorial analysis of nesting density, index of similarity of birds population on the model sites and structure of phytocenosis of the studied habitats.
The article presents topical preferences of several bird species which use succession biotopes in the exploited forests; key characteristics of the habitats transformed by the felling and determining the species composition; population density; ornithocenosis structure. We show the pattern of communities ordination in the exploited forests by the method of multidimensional non-metrical scaling. We traced the direction of successional, anthropodynamical changes in the communities towards the boreal communities under their almost absolute impossibility of reaching climax stages under the conditions of modern standards of forestry management.
According to our results, the working out of recommendations for forests treatment in the particular territories of the region is possible, taking the nesting of common and rare bird species into account.
PDF-версия на сайте cyberleninka.ru
© 2018 г. В.В. Гриднева